Le week-end du 6 et 7 mars 98, L. GARNERY, généticien au CNRS à Gif-sur-Yvette (France) était l’invité du CARI à Louvain-la-Neuve et des apiculteurs du Sud-Hainaut et du Nord de la France à Le Quesnoy, entre Valenciennes et Maubeuge : deux lieux de réunion, mais un seul conférencier et surtout une seule passion, celle de l’abeille et plus spécialement pour beaucoup de participants, celle de l’abeille noire. L. GARNERY, en effet, est un spécialiste de la génétique de l’abeille et ses travaux l’ont amené à étudier les populations d’abeilles noires du Sud-Hainaut et de l’Avesnois. Sa conférence a enthousiasmé plus d’un participants; en voici un bref compte-rendu.
Le modèle abeille
Une question vient naturellement à l’esprit : pourquoi un chercheur étudie-t-il l’abeille plutôt qu’un autre insecte ou un autre animal ? Tout simplement parce que l’abeille constitue un bon modèle pour les scientifiques : son étude permet de résoudre bien des questions d’intérêt général, mais aussi de mieux connaître un insecte important au plan économique ou écologique.
Un intérêt économique et écologique
L’abeille présente bien entendu un intérêt économique considérable : pensons aux produits de la ruche, mais aussi et surtout à la pollinisation des cultures. La pollinisation des plantes sauvages représente aussi une contribution inestimable. L’abeille est une espèce clé et tous les scientifiques s’accordent aujourd’hui pour dire que sa disparition entraînerait de graves problèmes pour la nature (et donc pour l’espèce humaine qui en fait intégralement partie !). Dans ce contexte, il est indispensable de conserver l’abeille, ce qui ne pourra se faire qu’en étudiant la biodiversité de cette espèce.
Un intérêt scientifique
Les chercheurs poursuivent aussi des objectifs qui ne trouvent pas directement une application concrète, mais qui conduisent à une meilleure compréhension du monde vivant. Ainsi, la biologie particulière de l’abeille convient bien à l’étude des mécanismes évolutifs : quand et comment une espèce apparaît-elle ? Comment a-t-elle évolué jusqu’aujourd’hui ? Selon quelles modalités a-t-elle colonisé son aire de distribution actuelle ?
Chez l’abeille, les scientifiques sont spécialement intéressés par quatre caractéristiques.
Une aire de répartition très importante. L’abeille est présente depuis le cercle polaire jusqu’au cap en Afrique du Sud, et de l’Angleterre à l’Oural. Cette large répartition témoigne d’une capacité d’adaptation à des situations écologiques variées, et donc d’une grande variabilité biologique (biodiversité).
L’haplodiploïdie. Contrairement aux mammifères, les mâles d’abeille sont haploïdes et les femelles (ouvrières et reines) sont diploïdes. De même, il n’y a pas de chromosomes sexuels chez l’abeille, mais un seul gène du sexe. Lorsqu’une abeille est homozygote pour le gène du sexe, cela donne un mâle dont la larve est mangée par les ouvrières. Lorsqu’il y a beaucoup de mâles diploïdes, cela conduit à l’observation de couvain lacunaire.
L’haploïdie du mâle a aussi pour conséquence que les caractères qu’il porte s’expriment automatiquement (contrairement à un individu diploïde). De ce fait, les gènes délétères entraînent la mort du mâle : c’est une sélection draconienne qui ralentit la transmission de caractères nuisibles à la survie de l’espèce.
L’eusocialité. L’abeille vit en société perenne dans laquelle s’organise la division du travail. Très peu d’espèces ont adopté cette stratégie.
La fécondation multiple. Plusieurs mâles, environ une quinzaine dans nos régions, s’accouplent avec une seule reine (on parle de polyandrie). Ces mâles se rassemblent dans des congrégations, zones particulières où se déroulent les accouplements. Ce comportement est à la base d’un important brassage des gènes qui évite la consanguinité et augmente donc la diversité biologique.
Le genre Apis
L’étude de la biodiversité chez l’abeille commence pour L. GARNERY par l’étude des relations de parenté entre les quatre espèces d’abeilles sociales (il faudrait plutôt parler de quatre groupes d’espèces) qui composent le genre Apis: Apis mellifera, A. cerana, A. dorsata et A. florea.
Apis mellifera a une distribution séparée de celle des autres espèces (distribution allopatrique); celles-ci ont une aire partiellement commune (sympatrie), mais A. cerana remonte plus vers le Nord. Pour ces trois espèces à distribution asiatique, on observe un polymorphisme de taille (différences de la taille du corps) et l’occupation de niches écologiques différentes. De cette manière, les espèces n’entrent pas en compétition, mais exploitent chacune des ressources spécifiques de leur milieu.
Notons aussi que A. cerana et A. mellifera vivent sur plusieurs cadres et dans des cavités; ces deux espèces présentent aussi une aire de répartition plus septentrionale. A. florea et A. dorsata vivent à l’air libre sur un seul rayon. Leur aire de répartition est tropicale.
La phylogénie des espèces a été obtenue par l’analyse de l’ADN mitochondrial, technique sur laquelle nous reviendrons ci-dessous. Un ancêtre commun à toutes les espèces du genre Apis aurait vécu en Asie du Sud-Est il y a plus de 10 millions d’années, époque à laquelle on observe la divergence d’A. florea à partir de cet ancêtre. Apis dorsata a divergé un peu plus tard, il y a environ 9 millions d’années. La séparation d’A. mellifera et d’A. cerana est beaucoup plus récente et date seulement de 6 millions d’années.
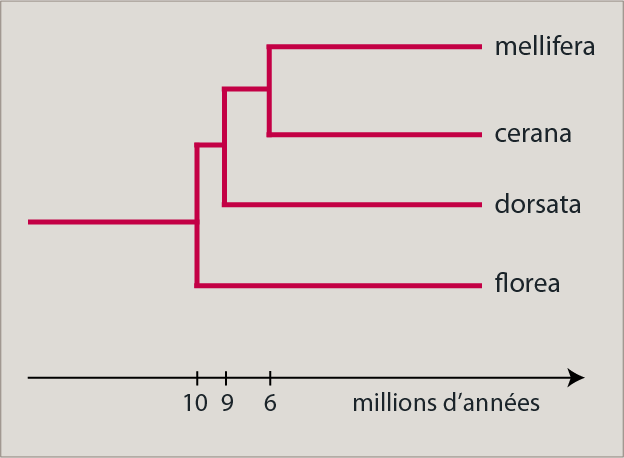
Comme on le voit, notre abeille est apparue très récemment (à l’échelle de l’évolution; l’âge de la terre est estimé à 4,5 milliards d’années). Pourtant, une tentative de colonisation de l’Europe a existé bien avant puisqu’on a trouvé un fossile d’abeille daté de 40 millions d’années en Allemagne. A cette époque, l’abeille avait donc déjà tenté de coloniser l’Europe, mais cette tentative s’est soldée par un échec.
Il faut aussi remarquer la congruence entre les données moléculaires et comportementales ou morphologiques : les espèces les plus apparentées, A. cerana et A. mellifera, sont aussi celles qui ont des comportements similaires (cavité, distribution septentrionale…).
L’espèce Apis mellifera
La variabilité au sein d’Apis mellifera est très importante. Différentes méthodes ont été utilisées pour l’appréhender. La plus connue des apiculteurs est certainement la morphométrie (appelée aussi biométrie).
La morphométrie
RUTTNER a beaucoup travaillé avec la biométrie. Il a mis en évidence l’existence de 23 à 25 sous-espèces ou races géographiques. Ces races se distinguent par une distribution géographique et un comportement particuliers. Chacun connaît par exemple, la distribution de l’abeille carniolienne (Yougoslavie, Autriche…) et ses caractéristiques comportementales (développement printanier précoce…). Tout cela est l’image d’une variabilité importante chez l’abeille mellifère.
Le polymorphisme enzymatique
L’étude du polymorphisme enzymatique (différentes formes d’une même enzyme) n’a pas été d’une grande utilité dans l’étude de la variabilité de l’abeille notamment à cause de la faible variabilité des marqueurs utilisés (il est difficile d’étudier la diversité avec quelque chose qui ne varie pas).
L’ADN
L’idéal pour étudier la variabilité consiste bien entendu à pouvoir étudier les gènes eux-mêmes puisqu’ils sont le support de cette variabilité. Avec les techniques actuelles de la génétique, c’est devenu chose possible. L’étude a d’abord porté sur l’ADN mitochondrial, puis sur l’ADN nucléaire.
ADN mitochondrial
Les mitochondries (organites cellulaires responsables de la production énergétique) possèdent de l’ADN qui « vit sa vie » de manière autonome; on parle d’hérédité extrachromosomique. L’ADN du mitochondrie est constitué d’une seule molécule circulaire que la reine transmet sans modification à tous ses descendants. L’abeille noire possède un ADN mitochondrial différent de celui des autre races. Le pouvoir de discrimination de l’ADN mitochondrial est très puissant puisqu’on peut reconnaître une colonie apparemment de race noire, mais qui a eu pour ancêtre une reine d’une autre race.
ADN nucléaire: séquences microsatellites
Les séquences microsatellites sont de courtes séquences d’ADN qui se répètent un grand nombre de fois et font preuve d’une grande variabilité, d’où leur intérêt pour l’étude de la biodiversité de l’abeille. Aujourd’hui, plus de 400 séquences microsatellites sont répertoriées.
Principes de base
Deux principes assez simples permettent de comprendre comment l’étude de ces molécules conduit à la connaissance de la phylogénie de l’abeille ou de la parenté entre colonies.
Principe de la ressemblance. La ressemblance entre deux fragments d’ADN est un signe de parenté, soit entre deux colonies (échelle de temps courte), soit entre deux espèces (aspect évolutif, échelle de temps longue). Prenons l’exemple de deux frères : le fait d’avoir des parents identiques implique que leur ADN est très semblable puisqu’il a une même origine.
Mise en évidence de la ressemblance. Les techniques de laboratoire permettent de mettre facilement en évidence les différences dans l’ADN. A l’aide d’enzymes de restriction – des « ciseaux » – on coupe la molécule d’ADN à étudier en fragments de différentes longueurs. La longueur de ces fragments varie en fonction de la variabilité de l’ADN : la taille des fragments chez deux colonies très apparentées est plus semblable que chez deux colonies non apparentées. Il suffit donc de séparer ces fragments et de les trier pour évaluer la ressemblance entre deux molécules d’ADN. Ce tri est réalisé par électrophorèse : le mélange d’ADN est déposé sur un gel soumis à un champ électrique. Les fragments, molécules chargées électriquement, migrent sur ce gel d’autant plus vite qu’ils sont courts. On assiste ainsi à une séparation des différentes tailles.
Étude de la variabilité par l’ADN mitochondrial
Trois lignées évolutives
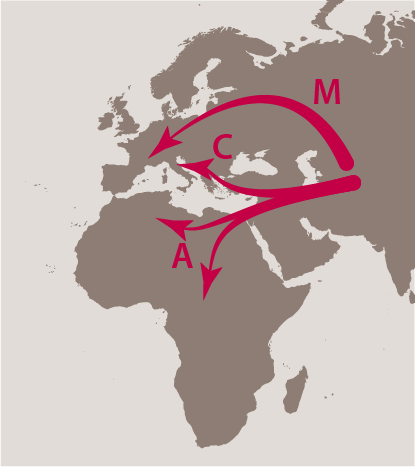
Un schéma clair se dégage de l’étude de la variabilité de l’ADN mitochondrial. Les colonies et les races se regroupent en trois lignées évolutives bien distinctes :
- la lignée africaine (lignée A) inclut tous les types correspondant aux races intermissa, adansonii, scutellata, capensis et monticola;
- la lignée ouest-méditerranéenne (lignée M) correspond aux colonies de l’abeille noire (A. mellifera mellifera et Apis m. iberica);
- la lignée nord-méditerranéenne (lignée C) inclut les races caucasica, ligustica ou carnica.
La distribution géographique des espèces actuelles du genre Apis conduit à penser qu’Apis mellifera est originaire d’Asie. De là, seraient parties trois grandes nappes (carte), l’une peuplant l’Europe par le nord (lignée M) en passant au nord des Alpes, du Caucase, de la mer Caspienne et Noire, l’autre longeant le nord de la Méditerranée (lignée C) et la troisième envahissant l’Afrique (lignée A). Une fois séparées par la distance ou les obstacles naturels, les trois populations ont divergé progressivement pour donner l’ensemble des races connues aujourd’hui. En se fondant sur le taux d’évolution de l’ADN mitochondrial, cet événement se serait produit entre -300000 et -1300000 ans, soit bien avant la dernière glaciation (-10000 ans).
La variabilité intralignée
La progression vers l’ouest de la lignée C peut être suivie facilement sur l’arbre phylogénétique. Les différentes populations s’organisent régulièrement depuis le Caucase jusqu’en Italie: il existe donc une structuration de cette lignée depuis l’Iran jusqu’en Europe.
Une telle variation n’existe pas dans la lignée M. Il faut y voir l’impact des glaciations. Au plus fort des périodes glaciaires, les abeilles de la lignée M se sont réfugiées dans le sud de la France et en Espagne pour recoloniser ensuite l’ensemble de l’aire actuelle. Cette réduction drastique de l’effectif d’une population se traduit généralement par une perte de diversité.
Le rôle des glaciations est moins marqué pour la lignée C qui dispose d’une aire de distribution plus méridionale et orientée est-ouest, alors que l’aire de répartition de la lignée M est plutôt orientée nord-sud.
Le plus intéressant pour l’abeille noire réside dans l’existence d’une sous-lignée typique du Sud-Hainaut en Belgique et de l’Avesnois en France. Le reste de la France est peuplé d’une autre sous-lignée (sauf l’Alsace qui est à rapprocher de la Belgique). Il faut donc envisager au moins deux conservatoires.
D’une manière générale, le taux d’introgression (les apiculteurs parlent de « pollution génétique ») de l’abeille noire est relativement faible dans les populations étudiées, ce qui est encourageant et conduit à penser qu’il est encore possible de prendre des mesures de conservation efficaces; mais il est grand temps !
Étude de la diversité par les séquence microsatellites
Parmi les nombreux résultats présentés par L. GARNERY, il faut certainement retenir deux choses. La première concerne la réelle originalité de l’abeille des populations Chimay et Valenciennes (soit le Sud-Hainaut et le nord de la France); ces deux populations (en fait, une seule population « à cheval » sur la frontière) sont vraiment différentes des populations du reste de la France. On est donc bien en présence de deux sous-lignées distinctes.
Le second point important concerne la diversité différente des trois lignées A, C et M. La variabilité de la lignée M est très faible, par rapport à celle des lignées C et A. Cela explique pourquoi la création de conservatoires est beaucoup plus urgente pour la lignée M (donc pour l’abeille noire) que pour les lignées A et C.
Perspectives
Ces travaux apportent bien des réponses aux questions que se posent les biologistes dans des domaines peu connus de la plupart des apiculteurs, notamment au niveau des mécanismes évolutifs et donc de l’histoire de notre abeille, depuis sa divergence à partir d’un ancêtre commun jusqu’aux quatre espèces d’abeilles du genre Apis.
Ces résultats scientifiques de base (on parle de recherche fondamentale) apparemment fort éloignés des préoccupations apicoles sont néanmoins le point de départ d’applications très utiles à l’apiculture.
- La biologie moléculaire apporte des outils nouveaux pour la reconnaissance des races d’abeilles. Aujourd’hui, les apiculteurs utilisent encore la biométrie. Cet outil a montré son utilité, mais aussi ses limites. Par exemple, l’utilisation aveugle de la biométrie sélectionne des colonies correspondant bien aux standards de la race, mais pas nécessairement aux caractères adaptatifs et zootechniques.
- La biologie moléculaire permettra aussi de reconnaître facilement les écotypes, populations locales ayant des adaptations spécifiques à l’environnement dans une région donnée.
- La connaissance des gènes liés aux caractéristiques zootechniques (douceur, production…) permettra de trouver très facilement les bonnes colonies dans un rucher et va « révolutionner » la sélection en apiculture.
- La connaissance des flux génétiques dans une population permettra de déterminer la taille de la population et la superficie nécessaire à la conservation; elle permettra de déterminer des marqueurs génétiques utiles au « pilotage » de la conservation de manière à éviter la perte de caractères intéressants.
- De manière plus anecdotique, la biologie moléculaire a montré que le sperme se mélange dans la spermathèque (en cours de vérification); elle permet aussi d’identifier dans la colonie les filles des différents mâles qui se sont accouplés avec la reine. Elle permet de même de connaître le nombre de mâles qui ont fécondé la reine.
- La biologie moléculaire permettra de déterminer l’impact des techniques apicoles sur les populations, par exemple la transhumance ou l’insémination artificielle (qui modifie la structure génétique de la colonie).
Conclusion
Ces nombreux résultats scientifiques apportent beaucoup d’informations nouvelles. Aujourd’hui les applications de terrain sont encore rares et il serait important que les associations apicoles s’organisent au niveau européen pour que ces techniques de laboratoire puisse être mises à la portée des apiculteurs dans le futur.
Par contre, sur le plan de la conservation des races d’abeilles, ces résultats apportent des arguments de poids en faveur de la conservation de l’abeille noire :
- les outils de la biologie moléculaire permettent d’établir de manière irréfutable la présence de l’abeille noire dans une région donnée;
- l’abeille noire existe encore dans de nombreuses régions, notamment en Belgique;
- les populations Chimay-Valenciennes sont tout à fait originales et seules représentantes d’une des deux sous-lignées connues à ce jour dans la lignée M;
- la création de conservatoires est urgente pour les populations de la lignée M, notamment de le sud du Hainaut et le nord de la France.